

Time to Tackle Traumatic Brain Injury
Time to Tackle Traumatic Brain Injury
Shan Lateef
Thomas Jefferson High School for Science and Technology
This paper was originally included in the 2019 print publication of the Teknos Science Journal.
Abstract
Traumatic brain injury (TBI) is a leading international cause of morbidity and mortality. The neurologic outcome of TBI is significantly influenced by the cellular and molecular responses (secondary injury) to the initial impact, which are poorly understood. The goals of this project were to: 1) Assess the level of apoptosis by immunolabeling cleaved Caspase-3 activity in whole Drosophila brains subjected to TBI, 2) Utilize quantitative real-time polymerase chain reaction, or qRT-PCR, and quantify the level of expression of 2 antimicrobial peptide genes (Attacin C and Diptericin B) after TBI, thereby assessing the role of the innate immune response, and 3) Detect the loss of intestinal barrier function in Drosophila, following TBI, by the presence of a non-absorbable blue food dye outside of the digestive tract, after feeding.
A “high-impact trauma” (HIT) device was built, which used a spring-based mechanism to propel flies against the wall of a vial, causing mechanical damage to the brain. An antibody to cleaved-Caspase-3 was used as a general apoptotic marker in whole Drosophila brains. The immune response was assessed by quantifying Anti-Microbial Peptides (AMP) gene expression using qRT-PCR. Loss of intestinal barrier function was detected by the presence of a non-absorbable blue food dye (FD&C blue dye no. 1) outside of the digestive tract after feeding.
Increased apoptosis was detected in Drosophila brains subjected to TBI by measuring cleaved caspase activity. TBI flies showed an enhanced innate immune response as measured by increased gene expression of the antimicrobial peptide, Diptericin B. More flies with TBI had the “Smurf” phenotype compared with controls, demonstrating that impaired gut permeability is a non-neuronal effect of severe TBI. TBI causes distinct biochemical and genetic alterations. An improved understanding of these secondary sub-cellular mechanisms of TBI is a vital prerequisite to developing effective interventions.
Introduction & Background
Traumatic brain injury (TBI) is a major public health concern and a leading international cause of morbidity and mortality [7]. Long term consequences of severe or recurrent TBI can include loss of balance, slower movements, tremors, and behavioral disturbances including mood swings, aggression, and even suicide [12]. Acute TBI is characterized by primary and secondary injury phases [15]. The primary brain injury is the direct mechanical trauma to the brain cells incurred at the time of the initial impact. This is followed by a series of biochemical events, within inflammatory, and cytotoxic pathways, which then result in secondary brain injury [21]. Thus, cellular and molecular responses to the initial impact significantly influence the eventual neurologic outcome of TBI. This delayed process of secondary injury allows hope for meaningful intervention. However, significant gaps still exist in our understanding of these secondary molecular mechanisms of traumatic brain injury.
Systematically assessing the impact of secondary injury pathways in mammalian models is difficult due to their relatively long life spans and high costs. Drosophila melanogaster has been used successfully to model human neurodegenerative disorders, and it was found that fundamental characteristics of human TBI also occur in flies [1, 9]. The fly and human brain have similar structural features and the fly cuticle, like the human cranium, is relatively inflexible and protects the fly brain from environmental insults. Furthermore, using flies has already provided novel insights into neurodegeneration, memory, and sleep, all of which are also affected in human TBI [3, 8, 11].
Phase 1 of this study created a successful animal model of single and recurrent TBI (rTBI) using Drosophila melanogaster and a HIT device [10]. Recurrent TBI had a measurable and significant negative impact on lifespan, time to sedate, and behaviors such as phototaxis (light affinity). Negative geotaxis (climbing) was not significantly affected by TBI. The most intriguing findings were that therapeutic hypothermia could significantly attenuate the deleterious effects of rTBI on lifespan, of mTBI and rTBI on time to complete sedation, and of rTBI on phototaxis [10]. The main goal of this study was to analyze the corresponding neuropathological changes, as well as the underlying cellular and molecular mechanisms, associated with these behavioral effects.
This project focused on 3 subcellular mechanisms of secondary injury:
Enhanced neuronal apoptosis: Apoptosis is a mode of programmed cell death which occurs normally during development in aging organisms and as a defense mechanism to eliminate imperfect or damaged cells TBI may activate apoptosis using a group of intracellular proteases called caspases, which are responsible for the deliberate disassembly of the cell. In fact, mouse models have implicated caspase-3 as a major mediator of apoptosis following TBI [6]. The activation of mammalian caspase-3 occurs after proteolytic cleavage, and this active form of caspase-3 can be recognized by a commercially available antibody. This antibody has also been shown to detect Drosophila effector caspases and was used in this study to qualitatively demonstrate apoptotic cells in the fly brains subject to TBI.
Upregulation of innate immunity: Secondary injury associated with TBI includes the rapid initiation of inflammatory pathways, which are necessary to initiate host defense responses and remove cellular debris [13]. However, the inability to appropriately attenuate these responses can lead to pathological inflammation that can further damage cells and tissues, thus worsening long-term outcomes [6]. Drosophila has a well-regulated innate immune system that uses antimicrobial peptides (AMPs) as effector molecules, including several with clear mammalian orthologs. Upregulation of this innate immunity may play a role in the neurodegeneration associated with TBI and this project specifically examined the role of two AMPs including Attacin C and Diptericin B, after rTBI, by quantifying the expression of these genes using quantitative real-time polymerase chain reaction (qRT-PCR).
Increased intestinal permeability (non-neuronal): Intestinal barrier dysfunction has linked metabolic and inflammatory markers of aging to death in Drosophila and may occur after head injury as well. Evidence from humans, rodents, and flies indicates that TBI can disrupt the intestinal barrier that normally functions to block the flow of certain ions, solutes, proteins, bacteria, and bacterial products between the inside and outside of the intestine [9]. Intestinal integrity can be assessed by the distribution of a blue food dye throughout the hemolymph post-feeding, and was employed in this study [17].
One factor that has been shown to worsen the effects of TBI in humans is hyperthermia, or increased body temperature [4]. Hyperthermia increases metabolic expenditure, glutamate release, and inflammatory activity that may further exacerbate neuronal damage [20]. Theoretically, hypothermia should mitigate the toxic effects of trauma which include excitotoxicity, free radical-induced alterations, inflammatory events, and disruption of the blood-brain barrier [1]. Studies have been conducted to determine the utility of hypothermia in severe TBI patients, but with conflicting results [11]. If temperature control could improve TBI outcomes, then it is crucial that we study this intervention systematically, and using an animal model would be a first step in this major initiative. Drosophila, being ectotherms, cannot endogenously regulate body temperature and essentially take on the temperature of their environments if physiologically permissible [5]. Prior research has shown that hypothermia or cooling Drosophila to 17℃ has a protective effect on brain potassium homeostasis during repetitive anoxia [18]. Cold temperature has also been shown to improve survival in Drosophila models recurrent TBI as well as mitigate some of the TBI associated behavioral effects in Phase 1 of this research [10].
In this study, a validated model of TBI in Drosophila melanogaster was used to investigate the three mechanisms of secondary injury and also determine whether hypothermia could exert its beneficial effect by mitigating one of these secondary responses. The objectives were:
To assess the level of apoptosis by immunolabeling cleaved-Caspase-3 activity in whole Drosophila brains subjected to TBI.
To utilize qRT-PCR and quantify the level of expression of 2 antimicrobial peptides (AMP) genes (Attacin C and Diptericin B) after TBI, thereby assessing the role of the innate immune response.
To detect the loss of intestinal barrier function in Drosophila, following TBI, by the presence of a non-absorbable blue food dye outside the digestive tract, after feeding.
Materials & Methods
Flies were obtained from Carolina Biological Company (wild-type, Oregon R strain Drosophila). Flies were maintained on formula 4-24® Instant Drosophila Blue Medium, also obtained from Carolina Biological, at 25 °C unless otherwise stated. As previously developed by Katzenberger and colleagues, a fly model of TBI was created by inflicting mechanical injury on flies using rapid acceleration and deceleration forces [9]. This produces outcomes that are similar to those of closed head TBI in humans. A “high-impact trauma” (HIT) device was constructed, consisting of a metal spring clamped at one end to a wooden board with the free end positioned over a polyurethane pad (Figure 1a and 1b). When the spring is deflected and released, the vial contacts the pad and the flies contact the vial and rebound, providing a model of TBI. The degree of injury can be adjusted by changing the extent of deflection or by varying the number of strikes.

Figure 1a. Inflicting TBI on Flies
Katzenberger, 2013

Figure 1b. Lateef HIT device
Lateef et al. 2019
Katzenberger and colleagues showed that deflection of the spring to 90° resulted in an impact velocity of ∼3.0 m/s (6.7 miles/h) and an average force of 2.5 N. Those flies subjected to a single strike with the spring deflected to 90° became temporarily incapacitated and fell to the bottom of the vial; however, there was no obvious external damage to the head, body, or appendages [9]. During the first minute after a strike, 8.8 ± 3.8% of flies were incapacitated, but most of these flies recovered locomotor activity within five minutes, as measured by climbing ability. Although mobility was reduced, it gradually returned over a two day period. The immediate loss of motor ability followed by ataxia and gradual recovery of mobility were reminiscent of concussion in humans and consistent with the idea that the HIT device inflicts brain injury in flies [9]. We observed a very similar response to single TBI in our experiment.
Traumatic Injury and Hypothermia: A deflection angle of 90 degrees was utilized and a total of 5 hits were applied, with one hit every 2 minutes. A one-day recovery period was allowed after the recurrent TBI, prior to starting the molecular assays. Hypothermia was induced for 3 minutes in a refrigerator at temperature of 16 °C, after all hits were performed. Moderate hypothermia in human trials was considered to be between 32- 34.5°C, which is approximately 1-5 °C lower than normal human body temperature. Therefore, our goal was to change the fly’s environmental temperature to 5-6 °C below its baseline. A one day recovery period was allowed after concussion or rTBI, before starting the behavioral assays. Katzenberger’s model of TBI shows that primary injuries cause death within 24 hours only if the injuries exceed a specific threshold. In addition, death from primary injuries is complete after 24 hours because the percent survival at 24 hours was not substantially different from the percent survival at 48 hours.
Apoptosis Detection Using Anti-Cleaved Caspase Antibody (Brain Dissection + Immunolabeling)
Flies were first anaesthetized with CO2, then beheaded and proboscis removed in 1x Phosphate Buffered Saline (PBS). Heads were then fixed in 4% paraformaldehyde for 45 minutes and brains carefully dissected. Drosophila brains were washed in 1x PBT (0.1% Tween 20 in 1× PBS ) three times for 15 minutes each at room temperature, then pre-incubated in fresh blocking solution for 15 minutes at room temperature on a rotating platform. Samples were incubated with cleaved-Caspase-3 antibody (1:100 dilution; Cell Signaling Technology) in fresh blocking solution, 2 nights at 4°C, on a rotating platform and washed again in PBT three times for 15 minutes each at room temperature. Samples were incubated with Alexa Fluor 568 (red) conjugated anti-rabbit antibody overnight at 4°C on a rotating platform. Another wash was completed in 1x PBT three times for 15 minutes each, followed by one wash in 1x PBS for 15 minutes. Samples were transferred to a glass slide with a drop of Vectashield and a coverslip was applied over the samples. Imaging was completed using a confocal microscope (Leica) with a fluorescent light source. Live and dead nuclei were visualized using DAPI (blue), or 4′,6-diamidino-2-phenylindole, and all Drosophila neurons were visualized using ELAV (Embryonic Lethal Abnormal Visual system), which is a green fluorophore. Cleaved-Caspase-3 was visualized using Alexa Fluor 568.
qRT-PCR to Quantify Gene Expression of AMPs
Ten flies were used per experimental group and were placed in a microcentrifuge tube with PBS no more than 1/3 of the volume. Flies were homogenized and mRNA was isolated using the RNeasy Mini Kit (Qiagen protocol). Then the Power SYBR Green RNA-to-Ct 1-Step Kit was followed for the gene expression assay (Thermofisher protocol). The Thermocycler ran with appropriate settings. PCR conditions were as follows: 40 cycles: step 1: 95 °C for 15 s, step 2: 60 °C for 60 s, step 3: 72 °C for 40 s each cycle. Primers were used for the genes: Attacin C, Diptericin B and dActin (reference).
Gut Reaction to TBI-Smurf Assay
This assay was used to determine intestinal integrity based upon the distribution of a blue food dye (FD&C blue dye #1) throughout the hemolymph, post-feeding. Briefly, three vials of 25 flies each were used for the 3 experimental conditions (control, rTBI, rTBI + cooling). After rTBI was inflicted, flies were transferred onto fresh medium containing blue dye (2.5% w/v) and allowed to feed overnight. Flies showing an extended blue coloration (not limited to the proboscis and crop) were considered “Smurf” flies, with disrupted intestinal permeability.
Results
A successful animal model of recurrent TBI was created using Drosophila melanogaster and the High Impact Trauma (HIT) device.
Detection of Apoptosis after Traumatic Brain Injury
The cleaved-Caspase-3 antibody was used to qualitatively demonstrate apoptotic cells in the fly brains (see Figure 2) which were co-labelled with DAPI and ELAV to demonstrate nuclei and neurons respectively. The injured brains demonstrated more neurons undergoing apoptosis than the control brains, mainly in the vicinity of the mushroom bodies. The group which underwent TBI and therapeutic hypothermia seemed to have less apoptosis than the group that was not cooled. This suggests that hypothermia may exert a beneficial effect after TBI by slowing down or inhibiting the enhanced apoptosis that occurs after brain injury.

Figure 2. Immunofluorescent staining to detect apoptosis in the Drosophila brain after TBI
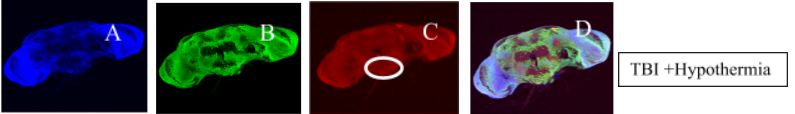
Figure 2. Drosophila brains co-stained with DAPI (A) which is a blue DNA-binding dye, ELAV (B) which is a green neuron-specific marker and anti-cleaved-Caspase 3 antibody (C) shown in red. Figure D in each panel shows the composite image with all 3 stains. TBI brains show more apoptotic foci (circled) and the TBI + hypothermia brains may show some attenuation of this apoptotic effect.
qRT-PCR Quantification of Anti-Microbial Genes After TBI
The relative quantification assay was executed after real time PCR amplification plots were analyzed. Rn is the fluorescence of the reporter dye divided by the fluorescence of a passive reference dye; ΔRn is Rn minus the baseline and is plotted against the PCR cycle number. Figure 3 represents the relative quantification of the genes across experimental groups. There was a significant upregulation of the AMP gene, Diptericin B, after TBI. Expression of the antimicrobial peptide (AMP) gene Diptericin B was significantly different across the 3 experimental groups by ANOVA (p <0.01). Post-hoc Tukey test showed that TBI and TBI + hypothermia flies had increased levels of Diptericin B compared to the control, and hypothermia did not diminish this effect. Unexpectedly, Attacin C expression did not increase among the injured flies, but did show an increase among flies subjected to TBI and hypothermia. There was no difference in dActin expression among the three experimental groups.
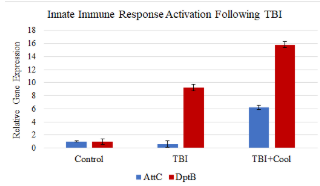
Figure 3. Relative levels of gene expression after TBI and with and without hypothermia
Disruption of Gut Permeability after TBI
After TBI, 13.3% of flies exhibited the smurf phenotype compared to controls at 0% (p <0.05). TBI followed by cooling caused 8.3% of flies to exhibit the smurf phenotype, which was less than TBI, but did not show statistical significance (p-value 0.5872).

Figure 4. Intestinal Integrity following TBI

Figure 5. Smurf fly after TBI
Discussion and Conclusion
A successful animal model of Traumatic Brain Injury was replicated using Drosophila melanogaster and a HIT device. This model was then used to study mechanisms of secondary brain injury and also to assess whether therapeutic hypothermia could mitigate any of these effects. Recurrent TBI caused enhanced apoptosis, activation of the immune response, and disrupted intestinal integrity. Therapeutic hypothermia seemed to have some effect at attenuating the enhanced apoptosis.
Immunostaining with an antibody against active Caspase-3 has been used to detect apoptotic cells mostly in Drosophila larval tissues. In this project it was successfully demonstrated that anti-cleaved-Caspase-3 antibody can detect apoptosis in Drosophila whole brain as well. This study confirms prior research in rats, mice and humans and highlights the importance of caspases in mediating sequelae of CNS injury [6]. This was a qualitative assay and precluded a finite assessment of apoptotic indices. The brain region that seemed to be most affected by apoptosis was close to the mushroom bodies that are known to play a role in olfactory learning and memory in Drosophila [16]. Apoptosis of neurons and glia contributes to the overall pathology of TBI, in both humans and animals, and can be caused by excitatory amino acids, increases in intracellular calcium, and free radicals [6]. Future studies may explore pharmacologic strategies that can reduce the extent of apoptotic cell death in animal models of TBI.
Injured flies exhibited a significant increase in the expression of the AMP gene, Diptericin B, which was not diminished by brain cooling. Attacin C levels were increased in the TBI + hypothermia group. These results indicate that the benefits of hypothermia [10] may not be lessened by damping down the inflammatory response. However, the TBI protocol used here inflicted severe TBI (5 hits in 10 minutes) as opposed to the single or recurrent TBI (4 hits in 48 hours) as employed in Phase 1 of this project. If inflammation is a significant part of the secondary injury cascade, combining anti-inflammatory agents with hypothermia may prove to be an effective strategy.
TBI disrupted intestinal barrier integrity as shown by the “Smurf” assay, where a blue dye dispersed through the entire fly body in the setting of intestinal barrier dysfunction [1]. Prior work has demonstrated that this increased permeability of the intestinal epithelial barrier results in hyperglycemia and a higher risk of death in flies [9], suggesting a possible role for dietary modification after severe TBI in humans.
This work builds upon my prior research and publication, which described a fly model of single and recurrent TBI that used a spring-loaded device to inflict closed-head TBI in adult flies. While Phase 1 of this research showed significant increase in mortality, a decrease in time to sedate, and impaired behavioral responses, this study attempted to focus deeply on the secondary Phase of injury which correlates with behavioral impairment and mortality as described in Phase 1. The TBI inflicted in this study was far more severe than in Phase 1, given that the primary goal was to elicit and characterize the secondary injury and it is possible that this is why hypothermia did not seem as effective. Future work should assess secondary injury for both mild and recurrent TBI separated by longer intervals to better represent human TBI.
Advances in the acute management of TBI are dependent on both an improved understanding of the metabolic, molecular, and cellular mechanisms that underlie neuronal and non-neuronal injuries, as well as the translation of such knowledge into the development of new molecules and techniques to improve the clinical outcome.
References
Peer Reviewed References
[1] Barekat, A., Gonzalez, A., Mauntz, E., Kotzebue et al. Using Drosophila as an integrated model to study mild repetitive traumatic brain injury. Nature Scientific Reports. 2016;6.
[2] Baxter SL, Allard DE, Crowl C and Sherwood NT. Dis Model Mech. 2014; 7(8):1005-1012.
[3] Bushey D, Cirelli C. From genetics to structure to function: Exploring sleep in Drosophila. Int Rev Neurobiol. 2011;99:213–244.
[4] Cairns CJ and Andrews PJ. Management of hyperthermia in traumatic brain injury. Current Opinion in Critical Care. 2002; 8(2): 106-110.
[5] Dillon ME, Wang G, Garrity PA and Huey RB. Review: thermal preference in Drosophila. J Therm Biol. 2009; 34(3): 109-119.
[6] Faden, A. I., Wu, J., Stoica, B. A. & Loane, D. J. Progressive inflammation-mediated neurodegeneration after traumatic brain or spinal cord injury. Br J Pharmacol, (2015).
[7] Johnson WD and Griswold WP. Traumatic Brain Injury: a global challenge. The Lancet Neurology. 2017;16 (12): 949-50.
[8] Kahsai L, Zars T. Learning and memory in Drosophila: Behavior, genetics, and neural systems. Int Rev Neurobiol. 2011;99:139–167.
[9] Katzenberger R, Loewen C, Wassermann D et al. A Drosophila model of closed head traumatic brain injury. PNAS. 2013;110(Suppl 44):4152-4159.
[10] Lateef, S., Holman, A., Carpenter, J., & James, J. (2019). Can Therapeutic Hypothermia Diminish the Impact of Traumatic Brain Injury in Drosophila melanogaster? Journal of Experimental Neuroscience. https://doi.org/10.1177/1179069518824852
[11] Lessing D, Bonini NM. Maintaining the brain: Insight into human neurodegeneration from Drosophila melanogaster mutants. Nat Rev Genet. 2009;10(Suppl 6):359–370.
[12] Ling H, Hardy J, Zetterberg H. Neurological consequences of traumatic brain injuries in sports. Molecular and Cellular Neuroscience. 2015;66:114-122.
[13] Lozano, D. et al. Neuroinflammatory responses to traumatic brain injury: etiology, clinical consequences, and therapeutic opportunities. Neuropsychiatr Dis Treat (2015) 11, 97–106, (2015).
[14] McKee AC, Cantu RC, Nowinski CJ et al. Chronic traumatic encephalopathy in athletes: progressive taupathy following repetitive head injury. J Neuropathol Exp Neurol. 2009;68(Suppl 7):709–735
[15] Park E, Bell JD, Baker AJ. Traumatic brain injury: Can the consequences be stopped? CMAJ 2008; 178(9):1163-70
[16] Rein, K., Zöckler, M., Mader, M., Grübel, C., & Heisenberg, M. (2002). The Drosophila Standard Brain. Current Biology, 12(3), 227-231. https://doi.org/ 10.1016/S0960-9822(02)00656-5
[17] Rera, M., Clark, R. I., & Walker, D. W. (2012). Intestinal barrier dysfunction links metabolic and inflammatory markers of aging to death in Drosophila. Proceedings of the National Academy of Sciences of the United States of America, 109(52), 21528-33.
[18] Rodriguez EC and Robertson RM. Protective effect of hypothermia on brain potassium homeostasis during repetitive anoxia in Drosophila melanogaster. Journal of Experimental Biology. 2012; 215: 4157-4165
[19] Tagge C, Fisher A, Minaeva O et al. Concussion, microvascular injury, and early tauopathy in young athletes after impact head injury and an impact concussion mouse model. Brain. 2018;141(Suppl 2):422-458.
[20] Thompson HJ, Tkacs NC, Saatman KE, Raghupathi R and McIntosh TK. Hyperthermia following traumatic brain injury: a critical evaluation. Neurobiology of Disease. 2003; 12(3): 163-173
[21] Veenith T, Goon SSH and Burnstein RM. Molecular mechanisms of traumatic brain injury: the missing link in management. World Journal of Emergency Surgery. 2009.
Non-Peer Reviewed




